
Evolution of Mammals Part 1 - Carboniferous to Cretaceous
Last updated: Tuesday October 19th, 2021
Report this blog
Introduction
The synapsids, a group including mammals and their ancestors, evolved during the late Carboniferous from basal amniotes. During the Permian, they were the dominant animals on Earth, before being supplanted in the Triassic by the archosaurs. The first mammaliaforms appeared in this period, having adapted to survive the archosaurs. Then, in the Jurassic, the first mammals appeared. They lived alongside giant reptiles, such as the dinosaurs and plesiosaurs, until the end of the Cretaceous, when an extinction event killed off three quarters of all life on Earth, including some of those early mammals. However, those that survived evolved to become the dominant animals on Earth. This blog consists of a list of animals from the Carboniferous to the Cretaceous which show the evolution of mammals and their ancestors, as well as animals with interesting features.
Archaeothyris
Archaeothyris was one of the two oldest confirmed synapsids, having evolved during the late Carboniferous. It was identified as a synapsid due to its skull: all synapsid skulls have a hole behind each eye, in a low position on the skull. Archaeothyris was more advanced than the reptiles alive at the time, having a strong jaw that it could open wider than its competitors. It was likely a carnivore, based on the teeth.

Ophiacodon
Ophiacodon was another primitive synapsid, which existed from the late Carboniferous to the early Permian. It was one of the largest tetrapods of its time, growing to between 1.6 and 3 metres in lenght and 26 to 230 kilograms in weight. It had a larger skull than other early synapsids, at 50 centimetres in length, and may have been partially warm-blooded.

Edaphosaurus
Edaphosaurus, also existing from the late Carboniferous to the early Permian, was one of the earliest-known large, herbivorous land vertebrates. Species of the genus reached between 0.5 and 3.5 metres in length and weighed over 300 kilograms. It had serrated teeth along the front and sides of its jaws, which it used to take bites out of plants, and teeth at the back of its mouth formed a surface that would crush and grind what the animal ate; it likely had powerful jaw muscles to help with this. As can be seen in the picture, Edaphosaurus had a sail on its back. Although related animals living at the same time, such as Dimetrodon, had sails, Edaphosaurus and other animals of the same family evolved sails independently. It is not known what the sail was used for.

Sphenacodon
Sphenacodon was a genus of synapsid belonging to the family Sphenacodontidae, which also included Dimetrodon. It was an apex predator, with the species S. ferocior growing as large as 3 metres in length. Like other members of its family, it had a crest along its back, though it was not as tall as the crests of animals such as Dimetrodon. It had three types of teeth: sharp pointed teeth which resembled incisors, large stabbing teeth resembling canines and small slicing teeth at the back. It lived during the late Carboniferous and early Permian.

Dimetrodon
Dimetrodon is one of the most famous synapsids, as well as one of the largest: species of the genus ranged in length from 60 centimetres to 4.6 metres and weighed between 28 and 250 kilograms. It lived during the early Permian and was likely an apex predator. Like Sphenacodon, it had incisor-like teeth, canine-like teeth and smaller teeth further back, and like other sphenacodontids the latter were teardrop-shaped. Different species of Dimetrodon had slightly different teeth: the teeth of smaller species lacked serrations because they hunted small prey, while the teeth of larger species had serrations, allowing them to slice through flesh. The most prominent feature of Dimetrodon was its sail, which may have been used to stabilise its spine, cool its body down or used in courtship display.
Although Dimetrodon is not a direct ancestor of mammals, it did have a feature that shows the transition between early tetrapods and mammals. In its nasal cavity were ridges called nasoturbinals, which may have supported cartilage. In later synapsids these ridges were larger, and may have supported mucous membranes that warmed and moistened air, suggesting warm-bloodedness.
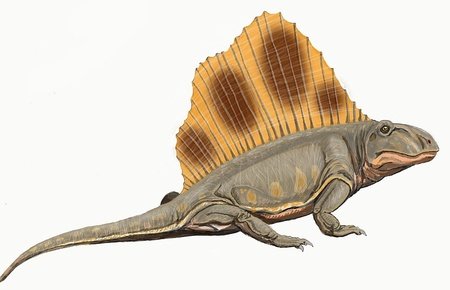
Biseridens
Therapsids, a group of synapsids represented today by mammals, first evolved during the Permian period from the group Sphenacodontia. Early therapsids were the first vertebrates to have limbs positioned underneath the body; they were also likely warm-blooded and probably had hair. Some therapsids had jaws that were more complex and powerful than those of other synapsids.
Biseridens was an early genus of therapsid, belonging to the clade anomodontia and alive during the middle Permian period. Like other anomodonts, it had a short snout compared to other therapsids, but it also had features different to those of other anomodonts such as multiple distinct teeth morphologies and denticles further back into the mouth.

Estemmenosuchus
Another group of early therapsids were the dinocephalians, which were dominant land animals of the middle Permian. One genus, Estemmenosuchus, grew to more than 3 metres in length and was an omnivore. It had large horns on the sides and top of its head, and was able to maintain an almost constant internal temperature due to its size and build.
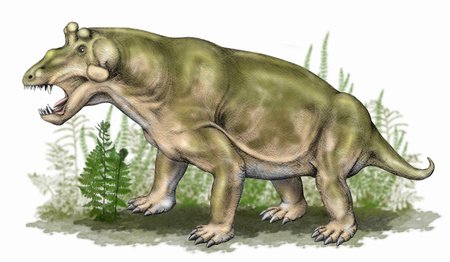
Anteosaurus
Anteosaurus was another large dinocephalian, growing to between 5 and 6 metres in length and weighing between 500 and 600 kilograms. It was, despite its size, a fast and agile predator with advanced senses of vision, balance and coordination, as well as large canines. Due to the thickness of its skull and its body shape, it has been suggested that it would head-butt rivals during contests or push prey to knock it over.
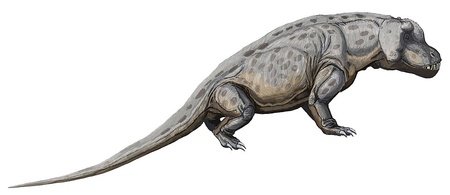
Moschops
Moschops was, like the previous two animals, a large dinocephalian, although unlike the other two it was a herbivore. It grew to 2.7 metres in length and had a large body, with a small head on a short neck. It had strong jaw muscles and stout teeth, suggesting that it fed on tough vegetation; as this food source would have been low in nutrients, Moschops likely spent most of its time feeding. Like Anteosaurus, it is thought that individuals competed with each other by head-butting.

Procynosuchus
The cynodonts are the group of therapsids that includes mammals, and they first evolved during the late Permian. Procynosuchus was one of the earliest cynodonts. It grew to 60 centimetres long and had features, such as a long tail and flat foot bones, that suggest that it may have been semi-aquatic.

Inostrancevia
The Gorgonopsians were therapsids which first evolved during the middle Permian and became extinct during the Permian-Triassic extinction event. After the dinocephalians became extinct, they became apex predators. Inostrancevia was the largest gorgonopsian, with species reaching 2.5 metres in length and weighing 300 kilograms on average. Like other gorgonopsians, they had massive canines, the ones in the upper jaw reaching 15 centimetres in length.
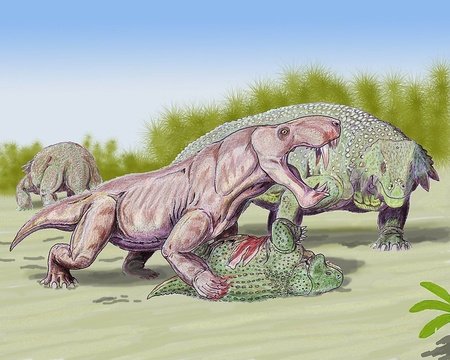
Moschorhinus
The therocephalians were a suborder of therapsids that lived from the middle Permian to the middle Triassic, and were the group most closely related to the cynodonts. They were carnivores, with some only growing to the size of rodents; however, Moschorhinus could grow as large as a lion. It also hunted in a similar way to a big cat, and had long incisors similar to the canines of sabre toothed cats, or to the gorgonopsians alive at the same time. Although Moschorhinus survived into the Triassic, individuals from this period had stunted growth.
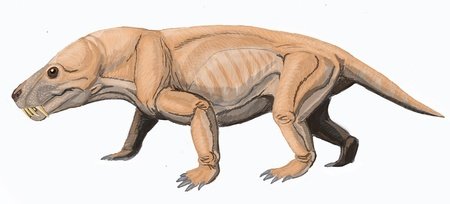
Diictodon
The dicynodonts first evolved during the middle Permian, and like the cynodonts they survived the Permian–Triassic extinction event. Like other dicynodonts, Diictodon had a pair of tusks and a beak, with the males having slightly larger tusks than the females. It was one of the most successful synapsids of the Permian period and was a burrowing animal, using highly developed muscles to dig burrows up to 1.5 metres deep. It lived in desert environments, which were common at the time, and nested near flood plains, meaning that occasionally the burrows would fill with water. Diictodon would use its beak to feed on desert shrubs, and it had a very efficient digestive system due to the lack of nutrients in these plants, similar to modern desert animals.
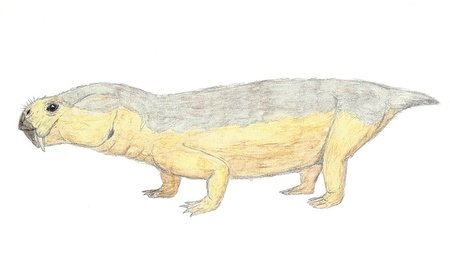
Endothiodon
There were larger dicynodonts than Diictodon: Endothiodon, which lived during the late Permian, could grow up to 2 metres long. Unlike most other dicynodonts, Endothiodon lacked tusks and had a distinct beak which likely allowed them to be specialised grazers. The teeth of juvenile Endothiodon were simpler than those of the adults, and more similar to those of a carnivore, suggesting that the young may have been insectivores or omnivores.
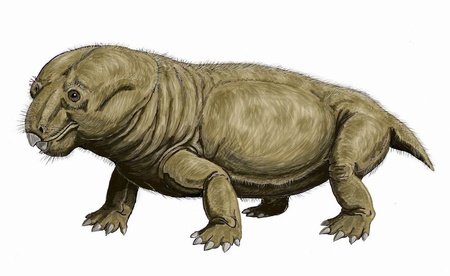
Lystrosaurus
Lystrosaurus was a genus of dicynodont that existed from the late Permian to the early Triassic. Species of Lystrosaurus grew to between 0.6 and 2.5 metres in length, and they had no teeth besides the pair of tusks. They dominated the southern part of Pangaea for millions of years during the early Triassic, to the point where 95% of the fossils in some fossil beds are Lystrosaurus. There are several theories as to why Lystrosaurus survived the Permian-Triassic extinction event, including that it may have been able to enter a state similar to hibernation or that its burrowing lifestyle made it able to cope with the increased carbon dioxide content in the air.
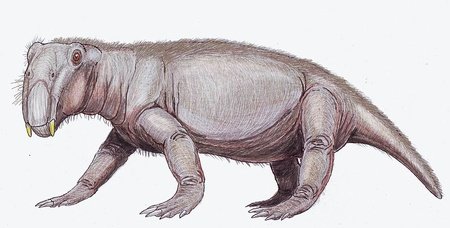
Thrinaxodon
Thrinaxodon was a genus of cynodonts that lived just after the Permian-Triassic extinction event. During this time, it was one of the few carnivores and, although it wasn't that large, it was larger than similar cynodont carnivores. One important adaptation that Thrinaxodon had was a secondary palate, which separated the nasal passages from the rest of the mouth: this would allow the animal to breathe when it had food in its mouth, allowing it to mash up its food more and as such decreasing the time it took to digest it. The skull of Thrinaxodon also had fewer bones than the skulls of earlier therapsids, making it more similar to a mammal's skull. However, unlike mammals, Thrinaxodon had a semi-sprawling posture that was partway between the posture of earlier synapsids and the upright position of modern mammals.
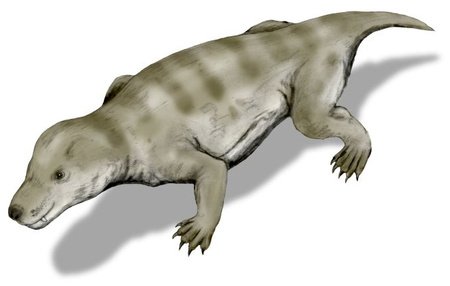
Cynognathus
Larger cynodonts had evolved by the middle Triassic, such as the 1 to 2 metre long Cynognathus. It had wide jaws with sharp teeth and a gait similar to some primitive modern mammals, with hindlimbs positioned directly beneath the body and forelimbs sprawled out like a reptile. It had a second palate like Thrinaxodon, but it may have also had an efficient diaphragm and whiskers.

Brasilitherium
Brasilitherium was an insectivorous cynodont alive during the late Triassic, and was small at approximately 12 centimetres in length and 20 grams in weight. It is an important animal, because it represents the transition between non-mammaliaform cynodonts and mammaliaforms. Its quadrate bone, which formed the lower jaw articulation in early synapsids, shows the beginning of the development of the stirrup (the bone in the middle ear). Impressions left on the skull by the brain suggest that Brasilitherium had a brain with early evolutionary traits of the mammalian brain, and it also had a tongue.

Lisowicia
Lisowicia, which lived during the late Triassic, was the largest known dicynodont and the largest non-mammalian synapsid to ever exist, weighing around 4 to 5 tonnes. It was also one of the last dicynodonts, as the dicynodonts died out by the end of the Triassic. It was unusual for a dicynodont, as its legs were positioned directly under its body; it also had no teeth, which includes the tusks found in most other dicynodonts. It mainly ate soft vegetation and conifers, but would occasionally eat tougher vegetation. Despite its size, it was hunted by some large predators, such as the archosaur Smok.
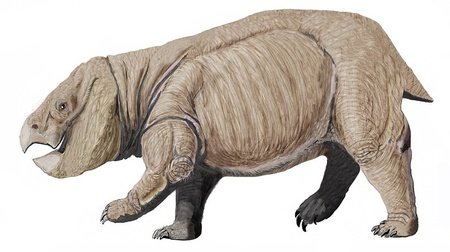
Morganucodon
The first mammaliaforms evolved during the late Triassic. This group contains the crown group mammals: the latter first evolved during the Jurassic, and earlier mammaliaforms such as Morganucodon are not considered to be true mammals. Compared to most other early mammaliaforms, many well-preserved fossils of Morganucodon have been found. It was a small nocturnal animal which spent the day in a burrow. It had glands used for grooming, suggesting that it had fur, and the fact that the young grew fast and had no teeth suggest that the juveniles were fed with milk, which may mean that Morganucodon was one of the first animals to lactate. Like mammals, Morganucodon individuals were born with primary teeth, which were then replaced with one more set of teeth that were not replaced when lost; and like monotremes, it is thought to have produced small, leathery eggs. Unlike mammals, it had a relatively slow metabolism, allowing it to live longer than a mammal its size would.

Pachygenelus
While most non-mammaliaform cynodonts became extinct by the end of the Triassic, two families that first appeared during the late Triassic survived. One of these was the family Tritheledontidae, which existed until the middle Jurassic. One genus was Pachygenelus, which existed during the early Jurassic. Despite the fact that it was not a mammaliaform, its jaw and enamel were similar to those seen in mammals.

Tritylodon
The other family of non-mammaliaform cynodonts to survive beyond the Triassic were the tritylodonts, which became extinct during the early Cretaceous. Tritylodon lived during the early Jurassic and grew to around 30 centimetres in length. They chewed their food, which likely consisted of plants, in a way similar to modern rodents. It was a burrowing animal, and gave birth to larval young like marsupials and monotremes do today.
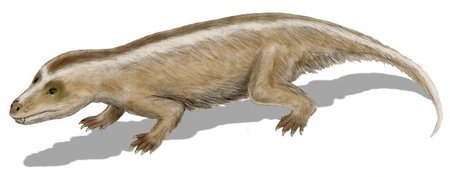
Gobiconodon
The first true mammals appeared during the middle Jurassic. Gobiconodon was a genus of mammal alive during this time, and was also one of the largest known mammals of the Mesozoic Era, growing up to 51 centimetres in length. It had shearing molars, large canine-like incisors and strong jaw muscles, suggesting that it was a predator.
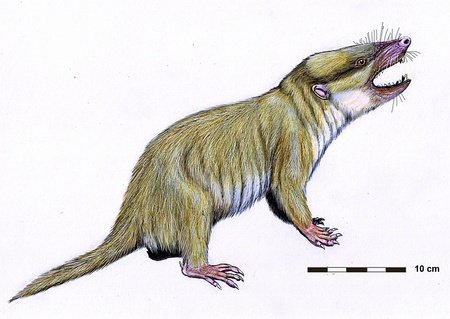
Agilodocodon
Other mammaliaforms, such as the docodonts, lived alongside the early mammals. One docodont alive during the middle Jurassic was Agilodocodon, which is the earliest known tree-climbing mammaliaform. It only measured 13 centimetres in length, which includes the tail, and weighed 27 grams. It had ankle and wrist joints similar to those of modern arboreal mammals.

Castorocauda
During the middle Jurassic, the mammaliaforms evolved to fill various habitats and niches. While Agilocodon was climbing trees, another genus of docodont, Castorocauda, was living a semi-aquatic lifestyle. Weighing between 500 and 800 grams, it was the largest known mammaliaform alive during the Jurassic. It is one of the earliest animals to have clear evidence of fur and had limbs adapted to either digging or swimming in a way similar to a platypus. It also had a spur on its hind ankle, like a platypus. However, it had teeth more similar to modern seals, suggesting that it ate fish.

Volaticotherium and Vilevolodon
These two animals have been included together because they both had the ability to glide. Volaticotherium lived slightly earlier, at 164 million years ago, making it one of the earliest-known gliding mammaliaforms. It was a mammal, and unusually for a gliding mammal it was carnivorous, though it likely fed on insects. Its gliding membrane not only extended between the limbs and tail base, but also between the digits, and its flat tail aided it when gliding.
Vilevolodon first appeared 161 million years ago, and was one of several related animals that could glide, including at least one genus which predated Volaticotherium. It, along with the other animals in its order, was either a mammal or a different type of mammaliaform. Unlike Volaticotherium it was likely a herbivore, largely feeding on seeds and soft vegetation, though it may also have been an omnivore. Unlike modern mammals, its ear bones were not entirely separate from the mandible.


Juramaia
The eutharians are a clade of mammals including the placentals but not the marsupials. Juramaia was one of the earliest known eutherians, living during the late Jurassic period. It was a small mammal, only growing up to 10 centimetres long, and its forelimbs have climbing adaptations, suggesting that the very first eutherians were arboreal.

Docofossor
Docofossor, alive during the late Jurassic period, was a docodont that was very similar to modern golden moles. It was around 9 centimetres long, had shovel-like fingers suitable for digging and a posture that suggests a subterranean lifestyle. Its upper molars were short and wide, something typical of mammals that forage underground.

Fruitafossor
Fruitafossor was a very small mammal, weighing only 6 grams, that lived during the late Jurassic. It likely fed on termites, like various modern mammals, based on its teeth: they were open-rooted and peg-like, and had no enamel, which is very similar to the teeth of armadillos and aardvarks. The forelimbs also support the idea that it ate termites, as they were large with powerful muscles, suggesting that they were used for digging.

Eomaia
Eomaia was a 10 centimetre long mammal alive during the early Cretaceous. Like Juramaia, it was more similar to placentals than marsupials, such as having an enlarged malleolus (a prominence at the bottom of the tibia) and similar jaws and teeth. However, unlike placentals, it also had a malleolus at the bottom of the fibula as well as a greater number of incisors and premolars.

Repenomamus
Repenomamus was a relatively large mammal from the early Cretaceous, the largest species growing nearly twice as long as Gobiconodon at 1 metre in length. It was a carnivore, feeding on other vertebrates; this included dinosaurs, as the skeletal remains of a juvenile Psittacosaurus were found in the stomach of one Repenomamus fossil. It may have hunted larger animals, or it may have scavenged. It either gave birth to underdeveloped young like marsupials or laid eggs like monotremes, as it had epipubic bones (bones that project forward from the pelvic bones, which prevent the torso from expanding and as such prevent the animal from giving birth to large young).

Steropodon
The first monotremes appeared during the Cretaceous, although related mammals existed as far back as the Jurassic. Steropodon is one of the oldest known monotremes, alive during the late Cretaceous, as well as one of the oldest mammals discovered in Australia. It grew to between 40 and 50 centimetres, and like platypuses it had a bill.
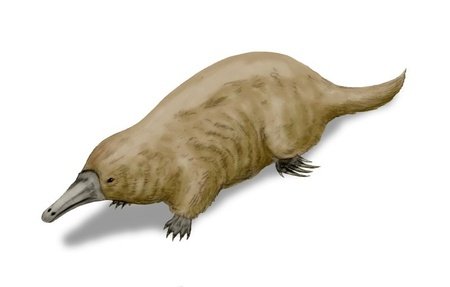
Alphadon
The metatherians, a clade consisting of marsupials and related extinct mammals, first appeared during the late Jurassic or early Cretaceous. Alphadon was a metatherian that lived during the late Cretaceous, which grew to around 30 centimetres in length. It was an omnivore with a powerful sense of smell and sight as well as whiskers, allowing it to track down its food during the day or the night.

Didelphodon
Didelphodon was another metatherian alive during the late Cretaceous, although unlike Alphadon it was a carnivore. The fact that it had large teeth and a powerful bite force meant that it was capable of biting through hard materials such as shells and bone; however, it likely fed on a mixture of hard foodstuffs and small vertebrates, as well as carrion.

Catopsbaatar
The multituberculates were the longest-living extinct mammal lineage, existing for 166 million years between the early Jurassic and the late Eocene. They were crown mammals, but were not therians or monotremes, although they were more closely related to the former. Catopsbaatar was a genus of multituberculates that appeared during the late Cretaceous. It had a wide skull, with small eye sockets and ridges that the jaw muscles attached to. The muscles themselves would have been strong, allowing the animal to break into hard seeds, although it also ate other animals. Like the platypus, Catopsbaatar had spurs on its hind legs, although there no evidence of a venom canal which can be found on the spurs of platypuses.

Vintana
The gondwanatheres were mammals that existed from the late Cretaceous to the early Miocene, which may have been related to the haramiyidans and the multituberculates. Vintana, which lived during the early Cretaceous, is known only from a single skull found in Madagascar. At a weight of 9 kilograms, it was one of the larger mammals alive during the Mesozoic Era.

Purgatorius
Purgatorius was a small eutherian mammal, only growing to 15 centimetres in length, which first evolved right at the end of the Cretaceous period and became extinct during the early Paleogene. It is thought to be the earliest known primate or proto-primate. It was an insectivore, and had ankles similar to modern primates, which suggest that it spend at least some of its time in the trees.

In Part 2
The next blog in this series will be focussed on the Paleogene. After most of the animals alive during the Cretaceous became extinct, mammals rapidly diversified to become the dominant animals on Earth.